
 Having demonstrated that nitrate increases pH on the root surface and ammonium reduces it, we will add that the influence of the two nitrogen ions is not limited to pH on the root surface, but indirectly affects, through the pH, the uptake of phosphorous, iron and all other elements the solubility of which is pH dependent (Fig. 4.21).
Having demonstrated that nitrate increases pH on the root surface and ammonium reduces it, we will add that the influence of the two nitrogen ions is not limited to pH on the root surface, but indirectly affects, through the pH, the uptake of phosphorous, iron and all other elements the solubility of which is pH dependent (Fig. 4.21).
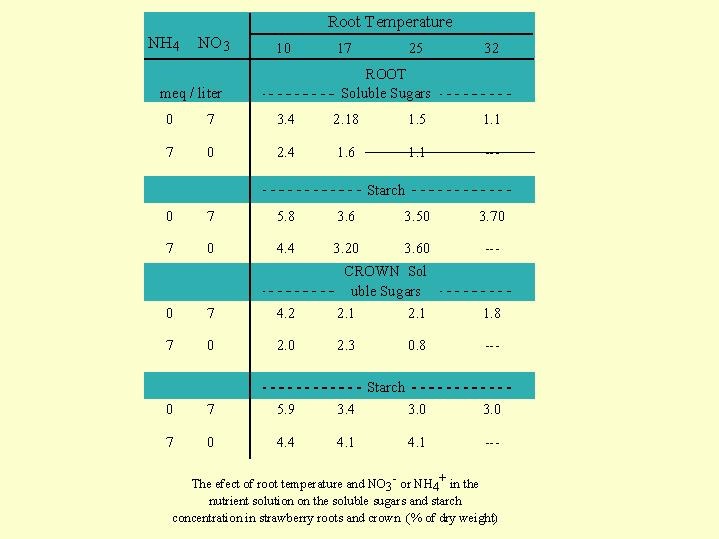
Another very important influence of the form of nitrogen in plant nutrition is shown in the experiment with strawberries in which all the plants were exposed to the same total N in the solution of 7 meq/L (98 gN/m3) but only the proportion of ammonium to nitrate varied (Fig 5.01).  Each NH4/NO3 ration was tested for 2 months at 4 different root temperatures: 10; 17; 25; 32 _C. At root temperature of 32_C and only ammonium in the solution the roots completely deteriorated, while at root temperatures of 10_C, also with only ammonium present, the roots and the plant was not damaged (Fig 5.01 D). The explanation to this behavior needs the understanding of nitrogen metabolism in the plant. Ammonium and nitrate are converted to amino acid in the plant according to the chemical equation in
Each NH4/NO3 ration was tested for 2 months at 4 different root temperatures: 10; 17; 25; 32 _C. At root temperature of 32_C and only ammonium in the solution the roots completely deteriorated, while at root temperatures of 10_C, also with only ammonium present, the roots and the plant was not damaged (Fig 5.01 D). The explanation to this behavior needs the understanding of nitrogen metabolism in the plant. Ammonium and nitrate are converted to amino acid in the plant according to the chemical equation in 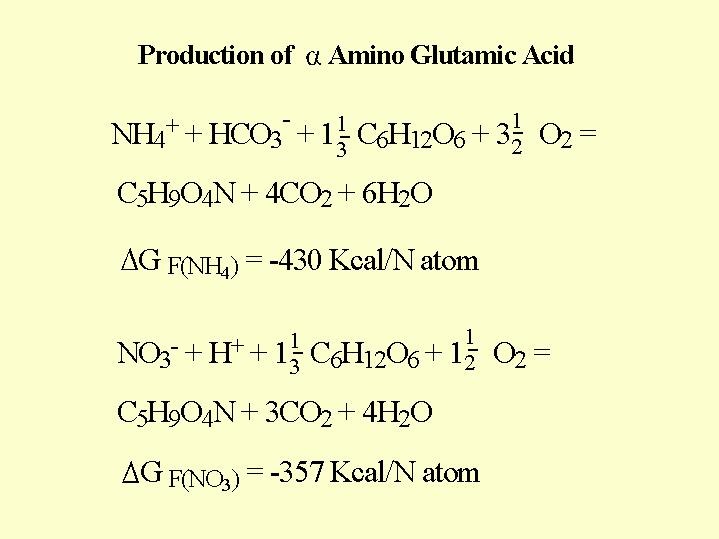 Fig. 5.06. Ammonium is metabolized in the root where it must meet the sugar that is delivered from the leaves. In the analysis of the strawberry plants it was found that there was a decline in the sugar concentration in the root cells as the ammonium content in the nutrient solution increased.(Fig. 5.02).
Fig. 5.06. Ammonium is metabolized in the root where it must meet the sugar that is delivered from the leaves. In the analysis of the strawberry plants it was found that there was a decline in the sugar concentration in the root cells as the ammonium content in the nutrient solution increased.(Fig. 5.02).
Free ammonium cannot stay as ammonium in the root not it moves upward as ammonium. In the cytoplasm the pH is above 7, which instantaneously will change the NH4+ partially into ammonia (NH3), which is very toxic to the respiration system in the cell. The ammonium is therefore immediately reacts with the sugar in the cell to produce amino acid. The nitrate is transported in the ionic form to the leaf where it is reduced to ammonium. In the leaf, the site of sugar production there is always excess of sugar as compared to the amonia produced during nitrate reduction. In the roots the situation is different. The sugar has to move from the leaves to the root. When the root temperature increases - the plant respiration also increases. As a result the sugar content in the root decline. The sugar is required simultaneously in large quantities for 2 reactions: 1-respiration and 2- ammonium metabolism. Therefore in winter, when the roots are cold, there is no problem in fertilizing with ammonium, whereas in summer (or when the roots are heated) - one should prevent using ammonium. This is critical especially for soil less systems in glass houses when the root containers are exposed to the sun and to the internal high temperature. The same high temperature that killed the ammonium fed plants did not harm the roots if only nitrate was present in the nutrient solution, ( Fig 5.01 A). The transport of sugars to the roots from the leaves is likely to lag behind it's consumption, particularly because the respiration and ammonium metabolism in the root continues day and night.
At a low root temperature, when the fertilization is only with nitrates, the root takes up the nitrate but it accumulates in the root since its upward movement to the leaves is restricted (  Fig. 5.03). The result is that not enough nitrogen will reach the leaves and the plant development is delayed. The practical result is that when the roots are cold ammonium nutrition is more appropriate. The actual "cold" or "hot" temperatures vary with each plant species and even cultivars.
Fig. 5.03). The result is that not enough nitrogen will reach the leaves and the plant development is delayed. The practical result is that when the roots are cold ammonium nutrition is more appropriate. The actual "cold" or "hot" temperatures vary with each plant species and even cultivars.
When glasshouse plants grew in clay pots the water evaporation from the clay produced a cooling effect and less damage to the roots was observed. Today, in nurseries with plastic pots, there is no evaporative cooling and as a result of the direct sun radiation the root temperature is increased and leads to greater sensitivity to excess ammonium in the fertilizer solution . In the work of Moritsugu et al. 1983, ( Fig 5.04), it is clearly seen that there is a difference in the sensitivity of various plants to the ammonium to nitrate ratio in the nutrient solution. Rice, barley, maize, sorghum, cucumber, tomato, lettuce, cabbage, Chinese cabbage, spinach, carrot and radish were given ammonium, nitrate or a mixture of ammonium and nitrate, with pH stabilization or without it, at a concentration of 5 mmol N (70 g N /m3).
Fig 5.04), it is clearly seen that there is a difference in the sensitivity of various plants to the ammonium to nitrate ratio in the nutrient solution. Rice, barley, maize, sorghum, cucumber, tomato, lettuce, cabbage, Chinese cabbage, spinach, carrot and radish were given ammonium, nitrate or a mixture of ammonium and nitrate, with pH stabilization or without it, at a concentration of 5 mmol N (70 g N /m3).
It was found that while rice and maize were relatively indifferent to the form of the nitrogen, Chinese cabbage and spinach plants died when they were given 5 mmol N as ammonium only (Fig. 5.04). The size of the root system and the sugar reserves in the roots explain the difference in the sensitivity of the various types of plants.
At a very low concentration, of 0.05 mmol N (0.70 g N /m3), Chinese cabbage lived well with ammonium nutrition, as the supply of sugars to the roots was sufficient for the N metabolism of this low level of ammonium (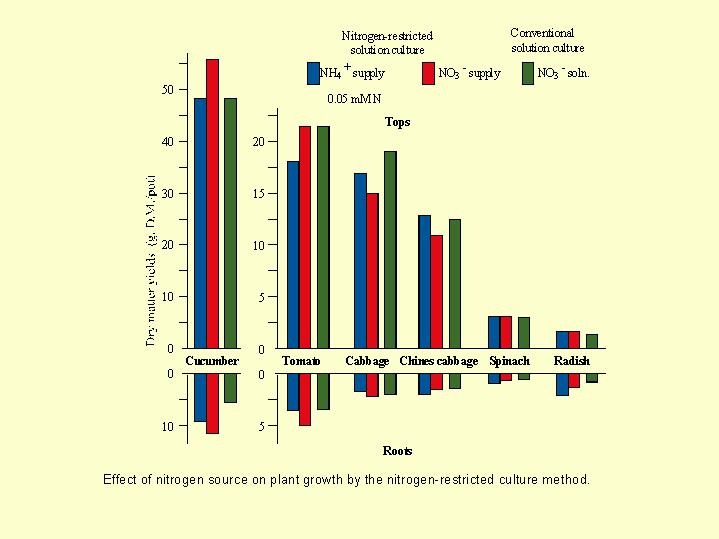 Fig. 5.05). Even the tomatoes lived well with ammonium at the very low concentration level but suffered decisive damage at 5 mmol N.
Fig. 5.05). Even the tomatoes lived well with ammonium at the very low concentration level but suffered decisive damage at 5 mmol N.
Accordingly ammonium should not be defined as "bad" or "good", but sophisticated usage should be made of the ammonium:nitrate ratio according to the growing conditions. In general it may be expected that crops, like Chinese cabbage, lettuce, spinach, with most of the growth is directed towards the leaves, will be more sensitive to high concentrations of ammonium than other plants that send large quantities of sugars to the roots.
A developing fruit is a strong sink for sugars, and reduces their availability to the roots. Accordingly, at the stage of fruit growth the plant will be sensitive to excess ammonium to a greater extent than in other periods.
With potatoes, during the tuber filling period, if there is insufficient phosphorous or potassium in the roots, the tuber will attracts them from the leaves and as a result the leaf blade will stop photosynthesis and carbohydrate supply to the tuber will also stop.
Calculating the free energy in production of one mole of alpha amino glutamic acid explains why the plant prefers the uptake of ammonium even when the "cocktail" contain both N forms (Fig 5.06). The plant gains about 80 kilo-calories for each gram atom of nitrogen in the ammonium form, as opposed to the production the same acid from nitrate.
On the other hand, the use of ammonium consumes much more oxygen. At higher temperatures less oxygen is dissolved in the water. When the roots are facing a "cocktail" containing ammonium and nitrate in a certain proportion the nitrogen form that will be taken up will vary with the temperature, kind of plant, and its growing stage. Plant growth depends on the type of the nitrogen, its concentration and the ratio between the ammonium and the nitrate and their effect on the carbohydrate content available in the plant.
An emerging seed , which is totally dependent on reserves from the seed and it's sugar reserves are limited, is more sensitive than the established plant to a high concentration of ammonium in its vicinity. A low concentration is tolerable but high concentration of ammonium can kill the developing seedling. An established cotton plant in the field with a large root volume will not die when ammonium or even ammonia fertilizer is placed near the plant even if partially of its root system is damaged. On the other hand a seedling near an ammonia band is likely to be destroyed immediately.
The metabolism cycle of the ammonium ( 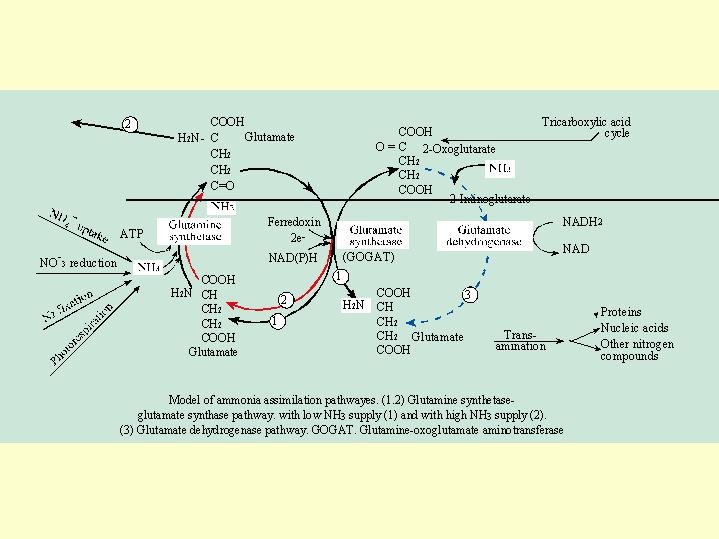 Fig. 5.07).
Fig. 5.07).
The cell cytoplasm contain ammonium from a number of sources:
ammonium uptake, reduction of nitrate, fixation of N2 from the atmosphere or nitrogen released in the photo respiration process.
The ammonia is immediately linked to the carboxylic group with the assistance of glutamate synthase and leads to the production of glutamine , which goes through a second cycle of glutamate production.
The important enzymes related to this process are glutamine synthase and glutamate dehydrogenase, which in their further action produce proteins, nucleic acids and the other compounds containing nitrogen, in the transamination processes.
At low levels of ammonia in the cell cycle No. 1 (Fig. 5.07) is operating producing glutamate (Only one -NH2 is bound to the carbon skeleton). When the load of ammonia in the cell increases cycle No. 2 is operating binding two -NH2 groups to the carbon skeleton.
The xylem solution transport to the top non-reduced nitrogen like nitrate, and reduced nitrogen. In old literature it was argued that there was no transport of nitrate in apples, in leaf analysis no nitrates were observed. There are plenty of evidence now that when the concentration of nitrates increases in the external medium to the roots - the nitrate accumulate in the top, while at low solution concentrations - there is nitrate reduction in the root and the nitrogen is transported in the xylem as reduced N compounds (Fig. 5.08).
{kind=link}
The amount of nitrogen reduced in the root to amino-compounds and moves in the xylem to the leaves varies among plant species. It can be seen (Fig.5.09) that the rate of reduction and N-metabolism in the root is greater with peas than maize or sunflowers. In peas nitrate and organic nitrogen are moving in the xylem at an almost equal rate. Some of the ammoniacal nitrogen stay in the top and some returns in the phloem to the roots. In maize only 25% of the nitrogen is transported in the xylem as organic and 2/3 as nitrates. 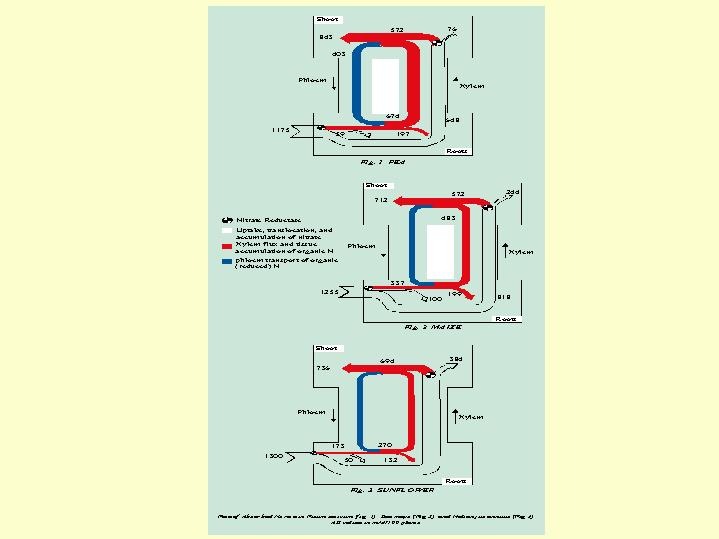 About 30% of the nitrogen returns in maize from the top to the root. On the other hand, with sunflower, only about 15% of the nitrogen in the xylem is found as organic nitrogen, close to 85% of the nitrogen in the sunflower is transported to the leaves in the form of nitrate. It's reduction occurs particularly in the leaves. Of course, a change in the concentration of nitrogen in the nutrient solution will change the ratios between the fraction of nitrogen reduced in the roots and that as nitrate in the xylem stream.
About 30% of the nitrogen returns in maize from the top to the root. On the other hand, with sunflower, only about 15% of the nitrogen in the xylem is found as organic nitrogen, close to 85% of the nitrogen in the sunflower is transported to the leaves in the form of nitrate. It's reduction occurs particularly in the leaves. Of course, a change in the concentration of nitrogen in the nutrient solution will change the ratios between the fraction of nitrogen reduced in the roots and that as nitrate in the xylem stream.
Potassium Role in the Plant
Leigh and Wyn Jones (1984) ( Fig. 5.10), described the relation between the potassium content in the dry matter and the crop as a % of the maximum yield.
Fig. 5.10), described the relation between the potassium content in the dry matter and the crop as a % of the maximum yield.
In most plants there is a steep increase in the yield up to a particular potassium content in the dry matter (1% in this example), and subsequently, although the potassium in the dry matter can increase, there is no additional response to the potassium in the form of yield increase. Accordingly the " critical K concentration" is defined as the concentration of potassium in the dry matter in plants that produced 90% of the maximum yield.
The critical potassium values accepted for example: 1% in tomato, 2.2% in sunflower (see table in  Fig. 5.11). In some crops it can be seen that sodium and magnesium can partially replace the potassium, thanks to their being analogous to them from the point of view of the osmotic influence in the vacuole but K can not be totally replaced.
Fig. 5.11). In some crops it can be seen that sodium and magnesium can partially replace the potassium, thanks to their being analogous to them from the point of view of the osmotic influence in the vacuole but K can not be totally replaced.
The maximum concentration of K in the dry matter varies between 3.5% to even 10%.
In  Fig. 5.12 the % of K in the dry matter is shown on the horizontal axis, and on the vertical - the % of nitrogen as protein of total nitrogen. Below 1% K, (potassium shortage conditions), there is a delay in the synthesis of the protein. When the potassium concentration increase above 1% in cereals - the addition of K no longer affects the production of the protein.
Fig. 5.12 the % of K in the dry matter is shown on the horizontal axis, and on the vertical - the % of nitrogen as protein of total nitrogen. Below 1% K, (potassium shortage conditions), there is a delay in the synthesis of the protein. When the potassium concentration increase above 1% in cereals - the addition of K no longer affects the production of the protein.
Leigh and Wyn Jones (1984) suggested to divide the potassium in the plant into two compartments: 1- the cytoplasm in which a more or less stable concentration of about 150 mM K is observed and 2- the vacuole in which a widely varying K concentration is allowed . The 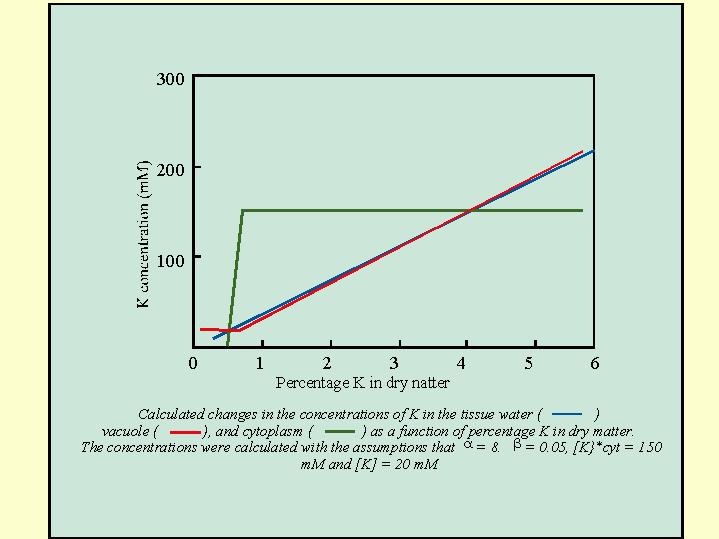 potassium begins to leave the cytoplasm only when its overall concentration in the dry matter drops below the critical threshold (about 1% in our example) and then there is a sharp drop in the cytoplasmic potassium (Fig. 5.13).
potassium begins to leave the cytoplasm only when its overall concentration in the dry matter drops below the critical threshold (about 1% in our example) and then there is a sharp drop in the cytoplasmic potassium (Fig. 5.13).
In Fig. 5.14 , their calculated model demonstrate the concentrations of K in the cytoplasm and the vacuole and the yield of the dry mater as a function of the concentration of K in the dry material. It appears that there is in fact a reduction, but not a dramatic reduction in the yield with the reduction in the concentration of K in the vacuole, but a steep drop in the yield occurs with a reduction of K content in the cytoplasm.
Why is such a high concentration of potassium required in the cytoplasm (150 mM are almost 6000 mg K per liter)? One of reasons is that the  potassium determines the ionic strength of the cytoplasm. The ionic strength is what determines the structure and the stability of the proteins in the plasma, which are required to maintain the precise enzyme activity of all the enzymes in the cell. Only potassium ion with it's specific atomic size and it's hydration energy is appropriate to ensure physical and physico-chemical characteristics of the plasma that are appropriate for the enzymatic activity. As soon as the concentration of K drops below the critical value no longer the biosynthetic activity is maintained at the same efficiency. The protein synthesis and the activity of all the enzymes in the cell is disturbed. In the sunflower, any drop below 10% of potassium in the dry matter involves a drop in the production of dry material. Only when the concentration in the vacuole is equal to the concentration in the cytoplasm is the maximum yield obtained (Fig. 5.14).
potassium determines the ionic strength of the cytoplasm. The ionic strength is what determines the structure and the stability of the proteins in the plasma, which are required to maintain the precise enzyme activity of all the enzymes in the cell. Only potassium ion with it's specific atomic size and it's hydration energy is appropriate to ensure physical and physico-chemical characteristics of the plasma that are appropriate for the enzymatic activity. As soon as the concentration of K drops below the critical value no longer the biosynthetic activity is maintained at the same efficiency. The protein synthesis and the activity of all the enzymes in the cell is disturbed. In the sunflower, any drop below 10% of potassium in the dry matter involves a drop in the production of dry material. Only when the concentration in the vacuole is equal to the concentration in the cytoplasm is the maximum yield obtained (Fig. 5.14).
With the move to drip irrigation, only part of the soil volume is wet and contributes potassium to the plant. It is therefore no wonder that there are increasing reports on response to potassium (such as "rusting" in cotton). Soil tests before seeding seem to show a sufficient supply of potassium. However, only about 40% of the soil volume actually supply K to the plants. Petiole tests frequently miss their role as a monitoring mechanism as they do not identify a developing K shortage tat early stages. The rate of the development of potassium shortage is very quick.
In now a classical experiment, with tobacco plants (Benzioni et al 1971), the plants were grown in a full nutrient solution whose composition included potassium nitrate. The plants were transferred on a particular date to a calcium sulfate solution (gypsum) for a day or a few days, before being returned to the complete nutrient solution.
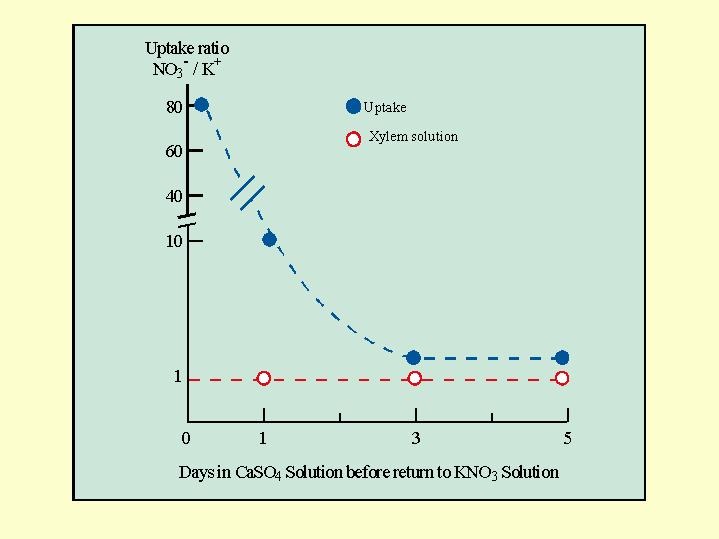 In the experiment the uptake of nitrate and potassium as well as their transport in the xylem were tested. The molar ratio of the NO3- to the K+ moving in the xylem on day 0 (in other words before the transfer of the plant to the gypsum solution) was 80 mole nitrate per 1 mole potassium. On the other hand, in plants that were returned to the full solution after 3 days of "nitrate starvation", when there were no more nitrates in the roots, the above ratio dropped to 1:1, which is similar to the ratio between these two ions in the nutrient solution (Fig. 5.15).
In the experiment the uptake of nitrate and potassium as well as their transport in the xylem were tested. The molar ratio of the NO3- to the K+ moving in the xylem on day 0 (in other words before the transfer of the plant to the gypsum solution) was 80 mole nitrate per 1 mole potassium. On the other hand, in plants that were returned to the full solution after 3 days of "nitrate starvation", when there were no more nitrates in the roots, the above ratio dropped to 1:1, which is similar to the ratio between these two ions in the nutrient solution (Fig. 5.15).
{kind=link}
In order to explain this system, the model of Benzioni, Vaadia and Lips, with the addition of Kirkby ( Fig. 5.16) was proposed. This model suggest that the nitrate anion accompanied with cations, during it's transport to the top. When the nitrate is reduced it's negative charge is transferred to the organic anions Malate, citrate, oxalate and pectate and their negative charge is balanced by the action K+ which is the major cation in the phloem stream, and partly by Mg2+ in their transport to the roots. Bicarbonate negative ion is released in the root to the solution and the root takes up nitrate anion from the solution. The potassium in the root that lost the bicarbonate, once again bind a new nitrate , and move with it to the top, unload the nitrate, load an organic anion, and moves down to the root, bicarbonate is released and so on.
Fig. 5.16) was proposed. This model suggest that the nitrate anion accompanied with cations, during it's transport to the top. When the nitrate is reduced it's negative charge is transferred to the organic anions Malate, citrate, oxalate and pectate and their negative charge is balanced by the action K+ which is the major cation in the phloem stream, and partly by Mg2+ in their transport to the roots. Bicarbonate negative ion is released in the root to the solution and the root takes up nitrate anion from the solution. The potassium in the root that lost the bicarbonate, once again bind a new nitrate , and move with it to the top, unload the nitrate, load an organic anion, and moves down to the root, bicarbonate is released and so on.
The calcium cannot replace potassium in this process since it back transport in the phloem is negligible. Calcium react in the vacuole with oxalate to produce precipitates of Ca-oxalate or with pectate to produced the stable Ca-pectate in the cell walls. Mainly the potassium (and to a lesser extent, the magnesium) fit for the transport role in the plant. In this way K makes about 1.5 cycles on the average, in the plant.
 In the case of continuous nitrate nutrition - the moles of potassium taken up is less than that of the nitrates absorbed.
In the case of continuous nitrate nutrition - the moles of potassium taken up is less than that of the nitrates absorbed.
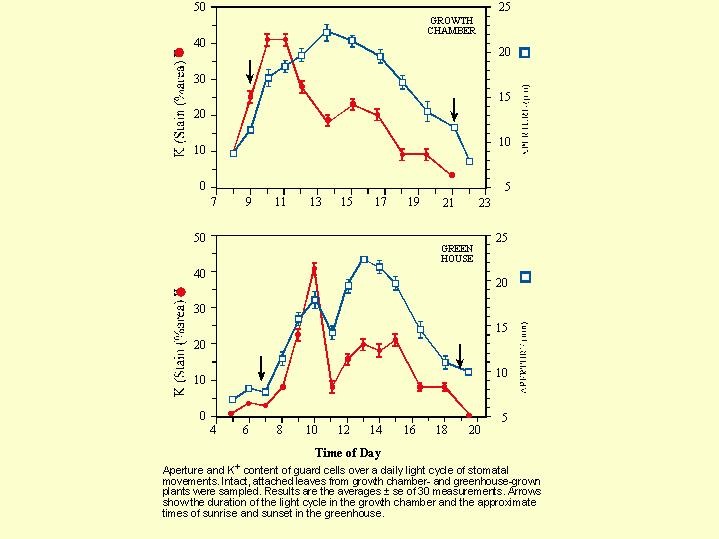
Talbot and Zeiger (1996) in a growth chamber and greenhouse experiments, describes the concentration of K+ in the guard cells of the stomata along the day and night (Fig. 5.17).
In the morning, at the beginning of the daily cycle, the concentration of potassium rose rapidly in the guard cells , the turgor increases and a short while thereafter the stomata opened. Later on at about 11 am the concentration of potassium in the guard cells decreased but 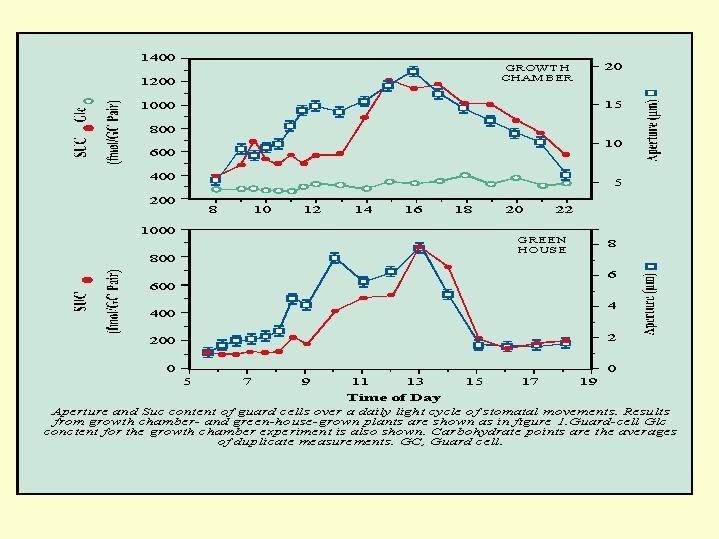 the aperture remained open for an appreciable period afterwards before eventual closing (Fig. 5.17).
the aperture remained open for an appreciable period afterwards before eventual closing (Fig. 5.17).
It appeared that for a period of time after the concentration of potassium decreased there was an increase in the concentration of sucrose, and only with the fading of the sucrose concentration, did the stomata closed (Fig. 5.18). The concentration of malate in the guard cells - rises with the potassium and drops almost simultaneously with the decline in K concentration of ( 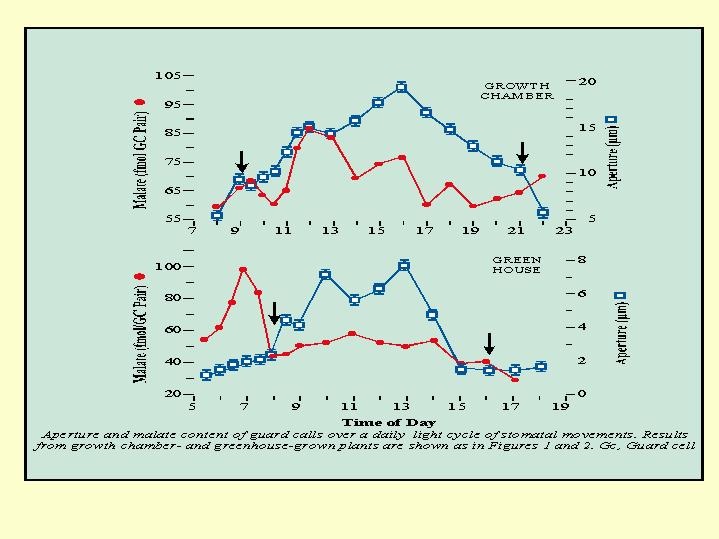 Fig. 5.19).
Fig. 5.19).
In the growth chamber no good link was found between the concentration of chloride in the guard cell and the opening of the aperture. A better relation was found in the greenhouse ( Fig. 5.20).
The concentrations of potassium and chloride in the aperture cells increase and decrease together and at very much the same rate. In other words in the stomata movements the chloride always accompany the potassium, in the guard cells of the stomata ( Fig. 5.21).
 Potassium Chloride constitutes the osmotic solution in the guard cells in the early morning hours before the sugar in the leaf is started to accumulate due to photosynthesis..
Potassium Chloride constitutes the osmotic solution in the guard cells in the early morning hours before the sugar in the leaf is started to accumulate due to photosynthesis..
In this experiment of Talbot and Zeiger the increase in the concentration of C02 in the greenhouse air led to the immediate closure of the apertures.
Upon returning to a normal C02 concentration, the apertures immediately re-opened. K leaving the guard cells accompanied the change in the concentration of C02 and the closing of the stomata (Fig. 5.22). Adding C02 at 1500h caused the closure of the apertures and the reduction in the concentration of sugar, but with no concomitant increase in the concentration of K in the guard cells. The reduction of the external air C02concentration caused the re-opening of the apertures. It appears that already by the afternoon K is lacking in the guard cells, and sugar replaces it as the osmotic controller of the stomata opening.. Although the light still continues, the apertures close, and the transpiration ceases, long before the light is switched off..
Why does potassium leave the guard cells in the afternoon? Why does sugar accumulate in its stead? And why as early as several hours before darkness is the concentration of sugar reduced and the apertures start to close? We do not yet have answers to offer to these questions. In practice, many measurements confirm that transpiration is reduced greatly in the afternoon.
The data in 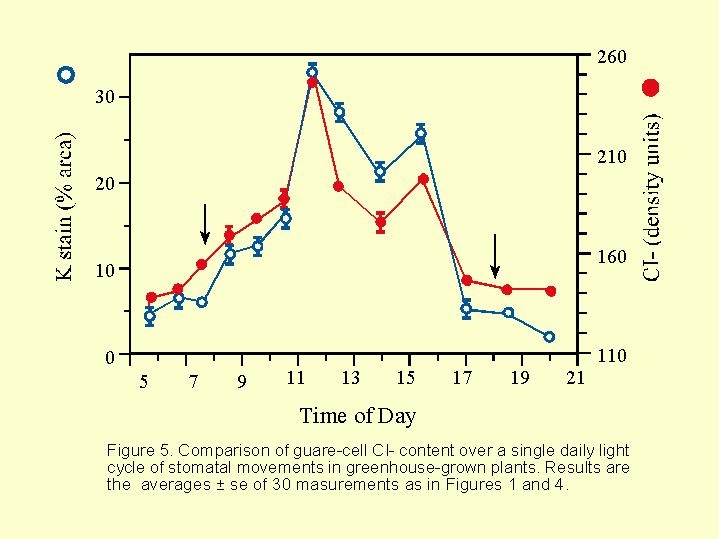 Fig. 5.21 seem to shed doubt the wisdom of adding C02 in greenhouses. This matter requires broad and detailed discussion in the light of this recent work of Talbot and Zeiger.
Fig. 5.21 seem to shed doubt the wisdom of adding C02 in greenhouses. This matter requires broad and detailed discussion in the light of this recent work of Talbot and Zeiger.